By Stephen J Gamble
Abstract
Higher life forms on Earth are multi-cellular organisms composed of eukaryotic cells. Eukaryotic cells are complex and are unlikely to have been the primary inhabitants of the planet. Evidence is examined to support the theory that eukaryotic cells evolved from simpler organisms by multiple endosymbiotic events where incorporated cells evolved into sub-cellular organelles. However, some researchers have proposed alternative mechanisms to the endosymbiotic theory for the presence of eukaryotic cells on Earth.
Introduction
In recent years there has been an increasing interest is in the search for life on other planets. For many centuries people on Earth have wondered about life elsewhere in the Universe, particularly intelligent life. At this point in time, the only place where life is definitely known to exist is on Earth, so the possibility and nature of life elsewhere is speculation. However, Davies (2007) quotes Belgian biochemist Christian de Duve as saying in 1995 that life is “a cosmic imperative” and that “it is almost bound to arise on any Earth-like planet”.
Understanding how life arose and then further developed on Earth might provide valuable indications on the possible nature of life elsewhere. Of course, completely different processes may be involved in the development of life elsewhere in the universe.
Although evolution is a continuous process the development to intelligent life can be spit down into a series of five major eras – the biochemical era followed by simple cells then complex cells, then multi-cellular organisms and finally intelligent life.
Pugsley (2018) identifies two key evolutionary issues that need to be resolved to allow the development of complex intelligent life. The first is homochirality – the handedness of molecules. For example, nearly all the amino acids used in organisms are the L-form and all the sugars are the D-form. Selecting molecules of the same handedness allow them to fit together to form chains. This needed resolution in the biochemical era before even simple cells could evolve.
The current discussion is restricted to the second of Pugsley’s evolutionary issues, the evolution of the other key aspect of life on Earth – the evolution of eukaryotic cells by endosymbiosis. This occurred as the third era of the evolutionary path described above and is a pre-requisite for the following two era. The events described here occurred either hundreds or thousands of millions of years ago.
What is endosymbiosis?
In classical symbiosis two organisms live together to their mutual advantage. The only difference with endosymbiosis is that one organism lives inside the cell of the second organism. The basis of the theory is that a number of sub-cellular components of eukaryotic cells arose from endosymbiotic arrangements where the incorporated cell became more integrated over time.
In looking at the origin of sub-cellular organelles this study will focus primarily on the origins of mitochondria and plastids. Plastids are found in higher plants and have a number of subtypes. The most widely known type of plastid is the chloroplast, a plastid containing the photosynthetic chlorophyll system.
Origins of the endosymbiotic theory
This is a theory that has been around for some time and has gained more support in recent years. Smith and Szathmary (2000) state (page 60) “In the early 1970s Lynn Margulis forcefully revived the idea of the symbiotic origin of plastids and mitochondria”, suggesting that the theory had been around before then. Gray et al (1999) state that the theory has been around for at least 100 years. There is a significant body of evidence to support this theory. Martin et al (2015), although mentioning earlier works that contain parts of the puzzle, attribute the first definitive endosymbiosis theory as being published in 1905 by Russian biologist Constantin Mereschkowsky.
Where do eukaryotes fit into the evolutionary tree
Life on Earth can be divided into three domains – the Bacteria, the Archea and the Eukaryotes. All higher organisms on Earth are complex multi-cellular Eukaryote organisms. Eukaryotic cells are primarily characterised by holding the majority of their genetic material separate from the general cell cytoplasm in a defined nucleus. They also have a number of other membrane bound sub-cellular organelles not seen in simpler organisms.
As a rough guide, 90% of organisms on Earth are either bacteria or archea. Of the remainder, around 99% are single cell eukaryotes (Walker, 2005, quoted by Gamble, 2008). This means that multi-cellular organisms make up only around 0.1% of all life on Earth.
When did it occur?
The Earth and the rest of the Solar system formed approx. 4500 million years ago. In addition to internal heating the Earth underwent severe meteoritic bombardment and did not form a stable surface until 3800 million years ago. The first evidence of living organisms (in the form of bacterial mats) appears around 3500 million years ago (Tritton,2001), although organisms may have existed before that date but have left no evidence.
Given the complexity of eukaryotic cells it is very unlikely that they were the first form of cell to evolve on Earth or originated at a similar time to bacterial cells. This paper will examine evidence describing how eukaryotic cells evolved from earlier simpler organisms. The common perception of evolution is that it is a straight-line process with one organism gradually changing into a new organism. In practice evolution is often a series of steps, some small, others large.
The first cellular organisms represent one of the large steps in evolution. Before that three other major steps must have occurred. Firstly, simple bio-chemicals must have formed from simpler chemicals. In a review of the synthesis of amino acids, purines and pyrimidines in primordial conditions, Miller (1998) states that this process occurred soon after the Earth’s formation. Formation of some of the chemicals may have taken place before the Earth had a stable surface, perhaps as a result of high energy from volcanos and massive lightning strikes as tested in Miller’s earlier experiments. However, Ranganayaki et al (1972) provide evidence that nucleic acid bases can be formed from simple chemicals under the action of sunlight in only a few days. The second major evolutionary step that occurred must have been the linking in someway of these simple bio-chemicals to make more complex molecules such as nucleic acids and peptides.
These first organisms were probably bacteria like and were prokaryotes. Prokaryote is a word derived from ancient Greek made up of pro, meaning before, and karyon, meaning a nut or a kernel. Basically, this means that a prokaryote is a cell without a kernel, or nucleus.
The formation of eukaryotic cells is itself another major step in the evolutionary ladder. Eukaryotic cells are much larger than prokaryotes. They have much more efficient ways to generate energy from nutrients using mitochondria whilst some have their own means to trap energy, for example, using chloroplasts. Mitochondria are thought to have evolved about 2200 million years ago whilst plastids start to appear about 1800 million years ago. This time line is not universally accepted, for example de Duve (2007) presents his view that eukaryotes appeared less than 1000 million years ago and quotes Cavalir-Smith as stating that the archea appeared at the same time as eukaryotes only 850 million years ago.
Poole and Penny (2006) present an alternative argument in which the Archea and the proto-eukaryotes arose independently at a similar time. Whilst their evidence supports the idea that the early eukaryote did engulf the mitochondrion they point out that there are no current examples of bacteria living in endosymbiotic relationship with archea.
These larger more efficient cells paved the way for the next big evolutionary step, the evolution of multi-cellular organisms containing a number of similar cells. It is a smaller evolutionary step from multi-cellular organisms to ones in which cells differentiate into specialised functions
The appearance of eukaryotic cells enabled the evolution of multi-cellular animals and plants. Tritton (2001) states that multi-cellular organisms originated around 600 million years ago. The evolution of multi-cellular organisms is, in turn, key to allowing the development complex intelligent life as we know it. The key step is the jump from prokaryotes to eukaryotes. Similar processes may, or may not, operate in other parts of the Universe.
How did it come about?
Roger (1999) states, “Resolving the order of events that occurred during the transition from prokaryotic to eukaryotic cells remains one of the greatest problems in cell evolution”. Compared to bacteria and the archea, eukaryotic cells are considerably more complex.
Giselle Walker (2005, quoted by Gamble 2008) stated that the most likely origin of eukaryotic cells is where one simple cell engulfed another cell. The engulfed cell was not digested but a symbiotic relationship was established. As the second cell is within the cell membrane of the primary cell this process is called endosymbiosis.
Walker supports the idea that structures such as mitochondria might have once been free living bacteria by pointing out that they have a double membrane. She postulates that the outer membrane could be derived from a vacuole formed by the host cell and the inner membrane could be derived from the cell membrane of the engulfed bacterium. There is further evidence that some early eukaryotes may themselves have been engulfed by other proto-eukaryotes as they have chloroplasts surrounded by a triple membrane. There may be evidence of a residual nucleus, called the nucleomorph, between the outer and middle membranes of the chloroplasts. So, chloroplasts may have already been a eukaryote cell (because they have a nucleus) before they were incorporated into another eukaryotic cell.
By the end of the 1980s it was thought that the general sequence of events was that the earliest organisms were the archea (originally the archea were thought to be a specialised branch of the bacteria called archeobacteria, whilst the true bacteria were termed eu-bacteria) which were followed by the eubacteria. Originally it was thought that the first eukaryote was formed when a eubacterium engulfed either another eubacterium or archeobacterium. However more recent work has shown the cytoplasmic structure and chemistry of archeobacteria to be more similar to that of the eukaryotes suggesting archea bacteria as the original host cells which engulfed other cells. This is supported by the findings of Pisani, Cotton and McInerney (2007) using complex phylogenetic analysis. For one cell to be able to engulf another cell the apparatus for phagocytosis (cytomembranes and the cytoskelton) must have been in place. De Duve (2007) states that if this is the case the evolution of the host cell towards being a eukaryote must have started long before the incorporation of the mitochondrial precursor. Margiluis et al (2007) confirm that this cytoskeletal motility must have occurred before mitochondria appear. As Martin et al point out, evolution by endosybiosis (as is horizontal gene transfer by way of plasmids) is at variance with classical Darwinian evolution by micromutation as favoured by de Duve.
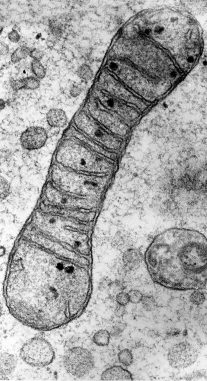
TM of mitochondria
credit: Prof. R. Bellairs CC BY
Both mitochondria and plastids have their own DNA. According to Kimbell (2005) this consists of a single circular molecule of DNA without any associated histone proteins. This is very similar to the prokaryotes and supports the idea that they were once free-living organisms. He further states that both mitochondria and plastids have their own protein synthesising machinery. The products of this synthesis all start with formyl methionine (fMet), as do proteins produced by bacteria, rather than methionine, which is the starting amino acid in proteins encoded by the DNA in the nucleus of eukaryotic cells. Following endosymbiosis, the incorporated bacteria that became sub-cellular organelles have reduced genomes with many of their genes being relocated to the nucleus of the host cell (Gray et al, 1999). Also supporting the idea that mitochondria and plastids were once free-living cell is that they both multiply within the eukaryotic cell by binary fission of existing mitochondria and plastids rather than being constructed from simpler proteins (Yoshida and Mogi, 2019).
The genomes of various members of the Rickettsiae family have been studied (Anderson et al 1998, McLeod et al 2004) and found to all be around 1.1Mbp in size. This is quite a small genome compared with a typical free-living bacterium such as E coli. Brzuszkiewicz et al (2011) studied two pathological strains of E coli and found the genomes to be around 5.31Mbp in size each. So, the Rickettsiae genome is only around 20% the size of other bacteria. Because Rikettsiae have reduced genomes they rely on the host cell for many functions, for example they do not have any of the enzymes required for glycolyis. They do have a complete set of enzymes for the TCA cycle and take up pyruvate as substrate from the host cell cytosol (McLeod et al).
Typically, mitochondrial DNA is around 16.5 kbp in size, just 1.5% of the size of the Rikettsiae genome, so they rely even more on either host genes or their ancestral genes transferred to the cell nucleus. Yang et al (2019) state that the mitochondrial genome codes of 2 ribosomal RNAs, 22 transfer RNAs and 13 polypeptide chains which combine with other subunits coded in the nucleus to form parts of the electron transfer chain. They state that more recently small hormones such as Humanin and MOTS-c have been found contained within the mitochondrial sequence for ribosomal RNA. These have effects on the local cell and in some cases more distant tissues.
If most of the genes required for the function of mitochondria and plasmids are stored in the cell nucleus there must be someway of co-ordinating between the two genomes. There has to be retrograde signalling from the mitochondria and plastids to the cell nucleus. Lee et al (2015) were able to show that MOTS-c is only encoded in the mitochondrial DNA and appears to have a regulatory function on transcribing some parts of the nuclear DNA.
Also supporting the endosymbiosis theory are the comments of Margulis and Sagan (2002). They state, “No missing links between eukaryotes and bacteria exist, either in the fossils or in life. The sudden appearance of eukaryotes on the evolutionary stage was genuinely discontinuous and not gradual”. However, a little bit of caution needs to be applied here – absence of evidence should not be used as evidence of absence. There may well be organisms in intermediate stages that have not yet been discovered or that previously existed and have died out. Other researchers (for example Poole and Penny 2006) suggest that the missing links, for example there being no existing examples of archea with a nucleus but no mitochondria, could be a problem with supporting the endosymbiosis theory. Since a eukaryote without mitochondria would be an anaerobic organism, many of which find high levels of oxygen toxic, it can be speculated that with the rise in oxygen in the atmosphere these relatively large organisms were unable to survive.
Martin et al (2015) mention organisms of the groups Guadia and Entamoeba have a nucleus but do not appear to have mitochondria. At first examination these would appear to be a missing link precursor to modern Eukaryotic cells with mitochondria. But it is believed that originally, they had mitochondria but lost them only being left with, what they describe as, relic organelles called mitosomes. So, these cannot be considered as one of the missing links.
An alternative, but not widely accepted, view could be that life on Earth arose because of panspermia could also account for missing links in the early evolutionary chain. The theory of panspernia, that life arose elsewhere in the universe and was transferred across space by interstellar dust, was first suggested in 1908 (Tritton, 2001). More recent work by Hoyle and Wickramasinghe (1979) suggests that interstellar dust grains could contain naturally freeze-dried bacteria and algae, and possibly viruses, and these could seed life on Earth. This would be consistent with de Duve’s 1995 comments highlighted above. There is no mention of the much larger eukaryotic cells being transferred by the mechanism. More controversially Francis Crick and Leslie Orgel (1973) state that levels of radiation in interstellar space are high enough to destroy known micro-organisms and they propose a mechanism of Directed Panspermia – where only selected micro-organisms are seeded on Earth by a extraterrestrial space craft. This could also account for missing links in the evolutionary history as only finished eukaryotes and not the early stages were selected.
The evidence seems to support the idea that engulfing followed by endosymbiosis has occurred at least twice in historic times to create eukaryotes as they are now observed. These engulfings seem to have occurred around 2200 million years ago and 1800 million years ago. Gray et al (1999) state that the genomes of mitochondria are sufficiently similar to support the idea that whatever organism mitochondria originated from was only engulfed at one time.
The story is further complicated by the discovery of hydrosomes which appear to be mitochondria like structures which produce hydrogen within cells. Originally, they were thought not to contain genetic material, but more recent studies have shown that they do. Gray (1999) postulates that hydrosomes, mitochondria and the eukaryotic nucleus may have arisen from a closely related group of eubacteria which invaded proto-eukaryotic cells at about the same time. This would suggest that engulfing has occurred, rather than just twice, at least four times in history which has resulted in different groups of sub-cellular organs.
Given that plastids have the remnants of their own circular DNA strand is suggestive that plastids were part of a later wave of endosymbiosis. Therefore, it is likely that hydrosomes, mitochondria and the nucleus were first incorporated into eukaryotes around 2200 milion years ago, whilst plastids were incorporated around 1800 million years ago. There is strong evidence (Bell (2001) and Forterre (2005, 2006)) that the eukaryote nucleus originated when a DNA virus invaded an early cell. Martin et alsuggest that both mitochondria and hydrosomes may have originated from a common ancestor organelle called perioxisomes. This could account for why mitochondria and hydrosomes appear to originate at a similar time.
Kimbell states that mitochondria are most closely related to aerobic bacteria, most probably the Rickettsias, whilst chloroplasts are evolved from cyanobacteria. In support of this he points out that a number of antibiotics such as streptomycin, which block protein synthesis in bacteria, also block protein synthesis in mitochondria and chloroplasts. Antibiotics do not block protein synthesis in the cytoplasm of eukaryotic cells. The antibiotic rifampicin blocks RNA polymerase in both mitochondria and eubacteria, but does not affect RNA polymerase in the nucleus of eukaryotes.
Conversely, Kimbell states that inhibitors of protein synthesis in eukaryotic ribosomes, e.g. diptheria toxin, have no effect on protein synthesis in eubacteria, mitochondria or chloroplasts. This suggests that bacteria, mitochondria and chloroplasts have a common origin.
It is interesting that Rickettsias bacteria, a group which are responsible for typhus and a range of tick-borne diseases, are suggested as the origin of these organelles as these are gram negative obligate intracellular parasites of eukaryotic cells. Given that they are normally intracellular parasites this might give an advantage in being able to enter a cell and forming a symbiotic relationship. Indeed, rather than an archeo-bacterium having engulfed a Rickettasia bacterium it is tempting to think that the archeo-bacterium was invaded. In practice Rickettsias bacteria attach themselves to the cell membrane of the host and await phagocytosis.
Conclusion
The situation can be summarised by a quote from Roger (1999) “Our current picture of early eukaryotic evolution is in a state of flux”. That situation still persists today as we find out more about the biology of both mitochondria and plastids.
The evidence presented primarily supports the theory that eukaryotic cells evolved from simpler organisms by incorporation of other simple cells, performing specialised functions, into a host cell by endosymbiosis. This process has occurred on multiple occasions in evolutionary history. The biochemical evidence supports the idea that a archea host engulfed or was invaded by, on multiple occasions, eubacteria which took on the role of subcellular organelles in eukaryotic cells. However, there is evidence that some of the structures necessary for phagocytosis and therefore also endosymbiosis to occur started to appear in the primordial eukaryotic cell as it evolved away from the other archea before the incorporation of these other cells.
Author
 Stephen J Gamble MIScT, FIBMS
Stephen J Gamble MIScT, FIBMS
Stephen has recently retired from a major research institute in the Cambridge area as a Research Assistant working on the molecular biology of cancers and ageing. As well as being a professional biologist, he has long standing interests in astronomy, astronautics and planetary science.
References
Andersson SGE, Zomorodipour A, Andersson JO, et al (1998) The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396, pp 133-140.
Bell PJL (2001) Viral Eukaryogenesis : was the Ancestor of the Nucleus a Complex DNA Virus? J. Molecular Evolution, 53, pp 251-256.
Brzuszkiewicz E, Thurmer A, Schuldes J et al (2011) Genome sequence analyses of two isolates from the recent Escherichia coli outbreak in Germany reveal the emergence of a new pathotype: Entero-Aggregative-Haemorrhagic Escherichia coli (EAHEC). Arch Microbiol 193, pp 883–891.
Crick FHC and Orgel LE (1973) Directed Panspermia. Icarus, 19, pp 341-346.
Davies P (2007) Are aliens among us? Scientific American 297, No 6, pp36-43
De Duve C (2007) The origin of eukaryotes a reappraisal. Nature Reviews Genetics, 8, pp 395-403.
Forterre P (2005) The two ages of the RNA world, and the transition to the DNA world: a story of viruses and cells. Biochimie, 87, pp 793-803.
Forterre P (2006) The origin of viruses and their possible roles in major evolutionary transitions. Virus Research, 117, pp 5-16.
Gamble, SJ (2008) The Evolution of Eukaryotic Cells. IST Journal Spring 2008 pp 8-9.
Hoyle F and Wickramasinghe C (1979) On the nature of interstellar grains. Astrophysics and Space Science, 66, pp 77-90.
Gray MW, Burger, G and Lang BF (1999) Mitochondrial Evolution. Science, 283, pp 1476-1481.
Kimbell, JW (2005) Endosymbiosis and the origin of eukayotes. Accessed online 5 March 2008
URL: http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/E/Endosymbiosis.html
Lee C, Zeng J, de Cabo R et al. (2015) The mitochondrial-derived peptide MOTS-c promotes metabolic homeostasis and reduces obesity and insulin resistance. Cell Metabolism 21, 443-454.
Margulis, L and Sagan, D (2002) Acquiring Genomes. Basic Books, New York.
Margulis L, Chapman M and Dolan MF (2007) Semes for analysis of evolution; de Duve’s peroxisomes and Meyer’s hydrogenases in the sulphouous proterozoic eon. Naturre Reviews Genetics, 8, pp 902-903.
Martin WF, Garg and Zimorski V (2015) Endosymbiotic theories for eukaryote origin. Philosophical Transactions of the Royal Society B 370, 20140330. Accessed online 3rd January 2018 at http://dx.doi.org/10.1098/rstb.2014.0330.
Miller, SL (1998) The endogenous synthesis of organic molecules. Chapter 3 IN: The Molecular Origin of Life (Ed: Brack, A), Cambridge University Press, Cambridge
McLeod M, Qin X, Karpathy S et al. (2004) Complete genome sequence of Rickettsiae typhi and comparison with sequences of other Rickettsiae. Journal of Bacteriology, 186, No 17, pp 5842-5855.
Pisani D, Cotton JA and McInerney (2007) Supertrees Disentangle the Chimerical Origin of Eukaryotic Genomes. Mo. Biol Evol 24 (8); 1752-1760.
Poole AM and Penny D (2006) Evaluating hypotheses for the origin of eukaryotes. Bioessays, 29, pp 74-84.
Pugsley D (2018) Do alien industrial civilisations exist? Journal of the British Interplanetary Society, 71, pp 443-449.
Ranganayaki S, Raina V and Bahadur (1972) Detection of nucleic acid bases in photochemically synthesized self-sustaining coacervates. Journal of the British Interplanetary Society, 25, pp279-285
Roger, AJ (1999) Reconstructing early events in eukaryotic evolution. American Naturalist, 154, Supplement 4 (Oct), pp S146-S163.
Smith, JM and Szathmary, E (2000) The Origins of Life. Oxford University Press, Oxford.
Tritton K (2001) Earth, Life and the Universe…exploring our cosmic ancestry. Curved Air Publications Ltd. ISBN 0-9540991-0-9.
Walker, G. (2005) Evolution of Eukaryotes. Lecture given 24 Feb 2005 at ACE, University of Cambridge, Homerton College.
Yang Y, Gao H, Zhou H et al (2019) The role of mitochondria-derived peptides in carcdiovascular disease: Recent updates. Biomedicine and Pharmacology, 117, ePub 109075.
Yoshida Y and Mogi Y (2019) How do plastids and mitochondria divide? Microscopy, 68, pp 45-56
Image
- TEM of a mitochondrion
- Credit: Prof. R. Bellairs CC BY